���̂��сA��ʌ��암�ō̏W�E�ώ@�����q���^�����f�[�^�����v100�̂ɒB�����̂ŁA
�����̕��͂������Ȃ�����Ă��������Ǝv���B
��������100�Ƃ����������ڕW�������킯�ł͂Ȃ��A�̏W��������荂�߂邽�߂ɁA��
�������̎���E�C���E�̒������G�N�Z���ɋL�^���n�߂��̂��[���ł��邪�A60�`70�̂ق�
�W�܂������_�ŁA���v100�̂ɂȂ�Γ��v�I�ɂ��L�ӂɂȂ邾�낤�Ƃ̍l���Ɏ������B
�����A�����܂Ŝ��ӓI�Ȋ����̌��ʂŁA�|�C���g�̑I����ώ@���Ԃ��s�K���ł���
���ƂȂǂ��l����ƁA���Ԋw�I�����Ƃ��Ă͕s�\���ł��邱�Ƃ�t�L���Ă��������B
�y100�̂܂ł̓��̂��z
�̏W�N�����Ƒ̒��i���̂݁j�A���Y�A����A���������ʒu�����\�Ɏ������B
��q����y�T�v�z�A�y�f�[�^���͕ҁz�̊�b�����ƂȂ��Ă���B
��2000�`01�N�̔������̏ڍׂ́A[�|�C���g�摜��]�q���^�N���K�^�̊ώ@�L�^���Q�Ƃ��������B
����ʎY�q���^�̕��ϑ̒��Ɋւ��ẮA�{�y�q���^���C���h���̒��f�[�^�i�s���{���ʁj���Q�Ƃ��������B
���̏W�̃q���g�Ɋւ��ẮA�q���^�N���K�^�̍̏W�Ƀ`�������W!!�@���Q�Ƃ��������B
| �v |
�N |
���� |
�̒� |
���Y |
���� |
�ʒu |
�v |
�N |
���� |
�̒� |
���Y |
���� |
�ʒu |
| 1 |
2000�N |
6��16�� |
48 |
�� |
�V���J�V |
���t |
51 |
2002�N |
8��28�� |
42 |
�� |
�N�k�M |
�� |
| 2 |
2000�N |
6��21�� |
30�` |
�� |
�V���J�V |
���� |
52 |
2002�N |
8��29�� |
34 |
�� |
�N�k�M |
���� |
| 3 |
2000�N |
6��26�� |
44 |
�� |
�V���J�V |
���� |
53 |
2002�N |
9��21�� |
35 |
�� |
�N�k�M |
�� |
| 4 |
2000�N |
6��27�� |
30 |
�� |
�V���J�V |
���t |
54 |
2003�N |
5��29�� |
37 |
�� |
�R�i�� |
�� |
| 5 |
2000�N |
7��3�� |
37 |
�� |
�R�i�� |
���t |
55 |
2003�N |
6��11�� |
���v�� |
�� |
�N�k�M |
�� |
| 6 |
2000�N |
7��10�� |
34 |
�� |
�R�i�� |
���t |
56 |
2003�N |
6��11�� |
���v�� |
�� |
�N�k�M |
�� |
| 7 |
2000�N |
7��16�� |
41 |
�� |
�N�k�M |
�� |
57 |
2003�N |
6��16�� |
42 |
�� |
�N�k�M |
�n�� |
| 8 |
2000�N |
7��19�� |
30 |
�� |
�V���J�V |
���� |
58 |
2003�N |
6��17�� |
49 |
�� |
�V���J�V |
���� |
| 9 |
2000�N |
7��21�� |
46 |
�� |
�R�i�� |
���� |
59 |
2003�N |
6��18�� |
44 |
�� |
�N�k�M |
�� |
| 10 |
2000�N |
7��22�� |
28 |
�� |
�R�i�� |
�� |
60 |
2003�N |
6��19�� |
50 |
�� |
�R�i�� |
�� |
| 11 |
2000�N |
7��26�� |
37 |
�� |
�N�k�M |
���t |
61 |
2003�N |
6��23�� |
46 |
�� |
�N�k�M |
�� |
| 12 |
2000�N |
7��28�� |
40 |
�� |
�N�k�M |
���t |
62 |
2003�N |
6��27�� |
���v�� |
�� |
�N�k�M |
�n�� |
| 13 |
2000�N |
7��30�� |
30 |
�� |
�N�k�M |
�� |
63 |
2003�N |
6��27�� |
46 |
�� |
�N�k�M |
�� |
| 14 |
2000�N |
7��30�� |
48 |
�� |
�N�k�M |
�� |
64 |
2003�N |
6��27�� |
���v�� |
�� |
�N�k�M |
�� |
| 15 |
2000�N |
8��7�� |
50 |
�� |
�N�k�M |
�n�� |
65 |
2003�N |
6��27�� |
38 |
�� |
�R�i�� |
���� |
| 16 |
2000�N |
8��7�� |
30�` |
�� |
�N�k�M |
�n�� |
66 |
2003�N |
6��27�� |
���v�� |
�� |
�V���J�V |
���� |
| 17 |
2000�N |
9��2�� |
30�` |
�� |
�N�k�M |
�� |
67 |
2003�N |
6��27�� |
32 |
�� |
�V���J�V |
���� |
| 18 |
2000�N |
9��2�� |
30�` |
�� |
�N�k�M |
�� |
68 |
2003�N |
6��30�� |
44 |
�� |
�N�k�M |
�� |
| 19 |
2001�N |
6��5�� |
50 |
�� |
�R�i�� |
���� |
69 |
2003�N |
7��4�� |
53 |
�� |
�N�k�M |
�� |
| 20 |
2001�N |
6��10�� |
38 |
�� |
�N�k�M |
�� |
70 |
2003�N |
7��10�� |
53 |
�� |
�N�k�M |
�� |
| 21 |
2001�N |
6��19�� |
47 |
�� |
�V���J�V |
���� |
71 |
2003�N |
7��22�� |
58 |
�� |
�N�k�M |
�� |
| 22 |
2001�N |
6��19�� |
���v�� |
�� |
�V���J�V |
���t |
72 |
2003�N |
7��24�� |
���v�� |
�� |
�V���J�V |
���� |
| 23 |
2001�N |
6��20�� |
48 |
�� |
�V���J�V |
���� |
73 |
2003�N |
7��31�� |
47 |
�� |
�N�k�M |
���� |
| 24 |
2001�N |
6��23�� |
40 |
�� |
�V���J�V |
���� |
74 |
2003�N |
8��1�� |
37 |
�� |
�N�k�M |
�� |
| 25 |
2001�N |
6��26�� |
50 |
�� |
�N�k�M |
���t |
75 |
2003�N |
8��4�� |
41 |
�� |
�N�k�M |
�� |
| 26 |
2001�N |
6��27�� |
36 |
�� |
�R�i�� |
�� |
76 |
2003�N |
8��20�� |
41 |
�� |
�N�k�M |
�� |
| 27 |
2001�N |
6��29�� |
40�` |
�� |
�N�k�M |
�� |
77 |
2003�N |
8��20�� |
38 |
�� |
�N�k�M |
�� |
| 28 |
2001�N |
6��30�� |
43 |
�� |
�N�k�M |
�� |
78 |
2003�N |
8��23�� |
37 |
�� |
�V���J�V |
���� |
| 29 |
2001�N |
6��30�� |
27 |
�� |
�N�k�M |
�� |
79 |
2003�N |
8��26�� |
34 |
�� |
�N�k�M |
�� |
| 30 |
2001�N |
6��30�� |
33 |
�� |
�N�k�M |
�� |
80 |
2003�N |
9��26�� |
30�` |
�� |
�N�k�M |
���t |
| 31 |
2001�N |
7��6�� |
41 |
�� |
�N�k�M |
�n�� |
81 |
2003�N |
9��26�� |
���v�� |
�� |
�N�k�M |
�� |
| 32 |
2001�N |
7��10�� |
36 |
�� |
�N�k�M |
���t |
82 |
2004�N |
5��17�� |
31 |
�� |
�V���J�V |
���� |
| 33 |
2001�N |
7��10�� |
41 |
�� |
�N�k�M |
���t |
83 |
2004�N |
5��21�� |
���v�� |
�� |
�V���J�V |
���� |
| 34 |
2001�N |
7��10�� |
53 |
�� |
�N�k�M |
�� |
84 |
2004�N |
5��29�� |
52 |
�� |
�N�k�M |
���t |
| 35 |
2001�N |
7��10�� |
58 |
�� |
�N�k�M |
�� |
85 |
2004�N |
6��10�� |
44 |
�� |
�R�i�� |
���t |
| 36 |
2001�N |
7��21�� |
43 |
�� |
�N�k�M |
���� |
86 |
2004�N |
6��14�� |
50 |
�� |
�V���J�V |
���t |
| 37 |
2001�N |
7��24�� |
39 |
�� |
�R�i�� |
�� |
87 |
2004�N |
6��17�� |
31 |
�� |
�N�k�M |
���t |
| 38 |
2001�N |
8��16�� |
58 |
�� |
�N�k�M |
�� |
88 |
2004�N |
6��22�� |
37 |
�� |
�N�k�M |
���t |
| 39 |
2002�N |
5��27�� |
47 |
�� |
�N�k�M |
���� |
89 |
2004�N |
6��22�� |
���v�� |
�� |
�N�k�M |
�n�� |
| 40 |
2002�N |
6��19�� |
37 |
�� |
�N�k�M |
�� |
90 |
2004�N |
6��22�� |
33 |
�� |
�N�k�M |
�� |
| 41 |
2002�N |
7��2�� |
40 |
�� |
�N�k�M |
���� |
91 |
2004�N |
6��22�� |
���v�� |
�� |
�V���J�V |
���t |
| 42 |
2002�N |
7��4�� |
39 |
�� |
�N�k�M |
�n�� |
92 |
2004�N |
6��23�� |
34 |
�� |
�V���J�V |
���� |
| 43 |
2002�N |
7��8�� |
48 |
�� |
�N�k�M |
�� |
93 |
2004�N |
6��29�� |
45 |
�� |
�R�i�� |
���t |
| 44 |
2002�N |
7��11�� |
27 |
�� |
�N�k�M |
�� |
94 |
2004�N |
6��29�� |
���v�� |
�� |
�R�i�� |
���t |
| 45 |
2002�N |
8��12�� |
40 |
�� |
�N�k�M |
�� |
95 |
2004�N |
6��29�� |
41 |
�� |
�N�k�M |
���t |
| 46 |
2002�N |
8��12�� |
34 |
�� |
�R�i�� |
�� |
96 |
2004�N |
6��30�� |
���v�� |
�� |
�N�k�M |
�� |
| 47 |
2002�N |
8��14�� |
41 |
�� |
�N�k�M |
�n�� |
97 |
2004�N |
7��5�� |
49 |
�� |
�N�k�M |
�� |
| 48 |
2002�N |
8��14�� |
33 |
�� |
�N�k�M |
�n�� |
98 |
2004�N |
7��12�� |
32 |
�� |
�N�k�M |
�� |
| 49 |
2002�N |
8��22�� |
51 |
�� |
�N�k�M |
�� |
99 |
2004�N |
7��14�� |
34 |
�� |
�N�k�M |
�� |
| 50 |
2002�N |
8��25�� |
���v�� |
�� |
�N�k�M |
�� |
100 |
2004�N |
7��23�� |
28 |
�� |
�N�k�M |
���� |
�y�T�v�z
�������n��
�����k���ōs����ꏊ���܂߁A���a��10�`�͈̔́B100�̂̂���99�͍�ʌ����Ŋm�F���ꂽ�́B1�̂�
�����s�������A��ʌ����ɂقNj߂��n�_�ł���A�����n����u��ʌ��암�v�ƈꊇ���邱�ƂɈّ��͂Ȃ��ł��낤�B
����������
2000�N6���`2004�N7���B���N5�`6���A�C���㏸�ɔ������������������A9�`10���ɉz�~���͂��߂�܂ł̊��Ԃɂ�����B
���t��̌`���Ə��łƂ����ω��̒��ŁA�ړ��ƒ蒅�A����ƎY�����s�Ȃ���B
���n�`�E�A��
�W����40�`180m�B
������u������̎G�ؗсv�ƌĂ�镽�n�сA���~�сA�u�˒n�̓сi�N�k�M�E�R�i����́j�ł���A�͐�~��
���i�M�͑ΏۂƂ��Ă��Ȃ��B���������āA��ʌ��암�₻�̎��ӂ̃q���^��������ԗ��������̂ł͂Ȃ��B
���̂̃J�E���g���@�Ƃ��̐��m��
2000�N�����唼���̏W�������A01�N�ȍ~�́A�ŏ��E�ő�̂ȂǕW�{�I���l������̂������A�̒��v�����
�����[�X���邱�ƂɓO�����B���������āA�z�~�̂��_�u���J�E���g���Ȃ��悤���ӂ����B�O�N�ɔ��������Ƃ����
�אڂ���ŗ��N�ɔ������ꂽ���T�C�Y�̌̂́A�f�[�^�Ɋ܂߂Ă��Ȃ��B
�������n��ւ̐l�דI�ȉe��
�`�F�b�N�����̂���9���͑��̍̏W�҂��K��Ă��邽�߁A�{���m�F�����ׂ��̐��́A���̃f�[�^�������Ă�
��͂��ł���B�Z��n�ɗאڂ����|�C���g�������A�Ǘ��҂ɂ�锰�́A��K�͊J���ɔ����G�ؗт̑r���ɂ��e����
�傫���B�������A�����̗v���������̐��̒����Ƃǂ�قǑ��֊W�����邩�ɂ��ẮA�ȒP�ȋc�_�ł͏I��
��Ȃ����낤�B�i��q�j
���u��ʓ암�Ńq���^100�́v�̋q�ϓI��Փx
�q���^�̏ꍇ5�V�[�Y����v�������A������G���A��100�̊m�F����܂łɗv���鎞�ԁi�\�z�j�́A�R�N���Ȃ�
1���A�m�R�M���Ȃ�1�`2�V�[�Y���ŏ\���ł��낤�B
�܂��A�����q���^�ő��n��Ɣ�ׂĂ݂�ƁA���ł�1�`2�V�[�Y�����H�@�܂��t�Ɋ֓��k���Ȃ�A��ʓ암���
�͂邩�ɍ���ɂȂ�Ǝv����B�[�I�Ɍ�����ʌ��암�́A�l�̈��D�ƃ��x���ł��������w�͂���A
�u�튴�o�̃q���^���̏W����B���������킦��n��v�ł͂Ȃ����낤���B
�y�f�[�^���͕ҁz
�@����ʍ̏W�̐�
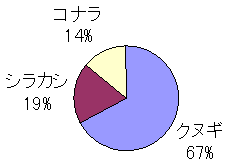
100�̂��m�F�������́A����3��ށB��ʂɂƂǂ܂炸�֓�����S�̂ōl���Ă��A�u���t�̏W�ɓK��������
��O�Ɓv�Ƃ����Ă��ߌ��ł͂Ȃ��B�Ƃ��ɃN�k�M�̓R�i���ɔ�ׂĎ��t�t���̌���⓴�������₷���A�i�D�̃z�X�g
�ƂȂ��Ă���B
�������n�̎G�ؗтɂ������ΐ��̓R�i�������|�I�ɑ������A��͂�|�C���g�̈��芴�A�m�������l����ƃN
�k�M���i���o�[�����ł���B�����A���w������ɍ̏W�����q���^�͂��ׂăR�i����������ł��������Ƃ��l����ƁA
�̏W���̗L����̐�����ɂ���ẮA�R�i�����N�k�M�Ɠ����̃p�t�H�[�}���X������\�����߂�
����Ƃ����悤�B���������̂̓V���J�V�B���t�̕��o�͋C�܂���ł����ʂł���ɂ�������炸�A�q���^���D��
�悤�ł���B�܂��A��L3��ȊO�ɂ����t���o������͂���A���Ӑ[���ώ@����Ύv��ʎ���ł����������҂ł�
��ł��낤�B
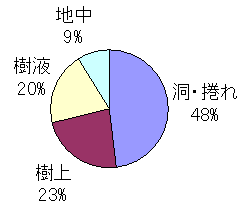
�A�����ꏊ
�������ɁA�������̋����N���K�^�̑�\�ł���B�n���Ɠ��E��������킹��ƁA57�����u�l�Ԃ��猩���Ȃ��ꏊ�v
�ɂ������ƂɂȂ�B�u����ł̔����v��23���ŁA�ꌩ�A��L�̐����Ɩ�������̂ł́H�@�ƌ�����ꂪ���ł���B
���A���������P�[�X�̂قƂ�ǂ́A�o����������̐V������z�~���������t��ֈړ��i���āj����O�i�K��
�������A�B��ꏊ�Ƃ��ēK��������⓴�����߂Ă���ق�̂킸���Ȏ��Ԃł�����B
���t�̏W�̎����ł���Ȃ���A���t��ő̂�I�o�����Ă���Ƃ���������̂�2���B
�悭�A�u�q���^��������Ȃ��������ɂ͂��Ȃ��v�ƒf�肷��������������邪�A����ɒ���t���Ă���J�u�g�A
�R�N���A�m�R�Ɠ��l�̐��Ԃ��q���^�ɑ��Ďv���`���Ă���ƁA��������襘H�ɂ͂܂�₷���̂ł͂Ȃ����낤���B
�������A�����������Ԃ�W�����Ō��J���邱�Ƃɂ���āA���������煂ȍ̏W�҂���������Ƃ�����A�r���⊶
�Ȃ��Ƃł�����̂����B
������c���t���o�Ă���ɂ��Ă��A���t�����1���ȏ㗣��Ă����ꍇ�́A
�@�@�@�@�@�@�u����v�Ƃ݂Ȃ��ăJ�E���g�����B
�B�����̔䗦
�u�Ȃ�����������Ȃ��̂��H�v
�q���^�̏W�ɂ�����ő�̋^��́A����ɐs����̂ł͂Ȃ����낤���B
�R�A�m�R�͂قځ��Ɠ����́������t��Ŋώ@�ł��A����������Ől�ڂ��͂��炸�i�H�j������ł��邱�Ƃ��قƂ��
�ł���B���ǂ���5�N�ԂŁA������̃q���^���O�Ō��邱�Ƃ͈�x���Ȃ������B�������߂�����Ńy�A������
�P�[�X�͎��X���邪�A���́��ȏ�Ɍx���S�������A�����Ő[���ɐ���ł��邱�Ƃ������B�܂��A�Y�������ɓ����
�����������t�̏W�͍���ɂȂ�B�܂��m���Ȃ͔̂����̎����B�����ĕp�x�͒Ⴂ���A�z�~�O��9���Ɉꕔ�̂���
�t��Ɏp��������Ƃ����Ō�̃`�����X�ł��낤�B
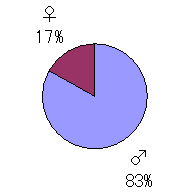
�C�������Ă������̌���
���̕\�́A�̂��m�F������������ʂɃi���o�����O�������̂ł���B�u�N�k�M�P�v�Ƃ́A�ŏ��Ɍ̂��m�F����
�ŁA�u�N�k�M27�v�͍ŋ߂Ɍ̂��m�F������\���Ă���B�܂��A���ꂼ��̖ŔN�x�ʂɊm�F���ꂽ�̐���
�����Ŏ����Ă���B���ۂɍ̏W�E�ώ@���ꂽ���̖{���́A�N�k�M��27�{�A�R�i��10�{�A�V���J�V5�{�̌v42�{�B
���̂����A�Ȃ�炩�̗v���ō̏W���s�\�������͒���������ɂȂ��Ă��܂������́A�v20�{�i�\�̐ԃA�~�����j��
�B���A�S�̖̂����߂Ă���B���N�A���������N���s���̂悤�ɐV�K�|�C���g��T������Ȃ��̂́A�܂���
���̂悤�ɂ��č̏W�ɓK�������������Ă�������ł���B
�u�̏W�ɕs�K�Ȏ��v�ɂȂ鎩�R�I�v���́A���Q�ɂ��|��A���̑S�ʂŎ��t�����o�������Ƃɂ��͊����͎�
�i�����͂�j�A�`���w�̐����ɂ�铴�̕��Ȃǂł���B�����ۂ��l�דI�v���ɂ́A��K�͊J���ɂ��|�C���g
���̂��̂̑r���A�̏W�҂ɂ����̔j���A�Ǘ��҂ɂ�锰�̂Ȃǂ�����B
|
�m�F���ꂽ�̐� |
|
| ���� |
00�N |
01�N |
02�N |
03�N |
04�N |
���̌���
�i�s�K���R�j |
| �N�k�M�P |
1 |
|
|
|
|
|
| �N�k�M�Q |
2 |
|
|
|
|
���� |
| �N�k�M�R |
2 |
|
|
|
|
|
| �N�k�M�S |
2 |
1 |
2 |
|
3 |
|
| �N�k�M�T |
1 |
|
|
|
|
�j�� |
| �N�k�M�U |
1 |
|
|
|
|
�j�� |
| �N�k�M�V |
|
2 |
|
|
|
�͊� |
| �N�k�M�W |
|
1 |
|
|
|
|
| �N�k�M�X |
|
5 |
1 |
|
|
�j���E�͊� |
| �N�k�M10 |
|
1 |
|
1 |
|
�͎� |
| �N�k�M11 |
|
2 |
|
|
|
|
| �N�k�M12 |
|
1 |
|
|
|
|
| �N�k�M13 |
|
|
3 |
2 |
2 |
|
| �N�k�M14 |
|
|
1 |
1 |
|
�|�� |
| �N�k�M15 |
|
|
2 |
1 |
|
|
| �N�k�M16 |
|
|
3 |
|
|
|
| �N�k�M17 |
|
|
1 |
|
|
�͊� |
| �N�k�M18 |
|
|
1 |
3 |
|
|
| �N�k�M19 |
|
|
|
3 |
1 |
�j�� |
| �N�k�M20 |
|
|
|
5 |
|
���� |
| �N�k�M21 |
|
|
|
1 |
|
�j�� |
| �N�k�M22 |
|
|
|
1 |
|
|
| �N�k�M23 |
|
|
|
2 |
|
�j�� |
| �N�k�M24 |
|
|
|
|
1 |
|
| �N�k�M25 |
|
|
|
|
2 |
|
| �N�k�M26 |
|
|
|
|
1 |
|
| �N�k�M27 |
|
|
|
|
1 |
|
| �R�i���P |
1 |
1 |
|
|
|
�j���E�͊� |
| �R�i���Q |
2 |
|
|
|
|
�|�� |
| �R�i���R |
1 |
|
|
|
|
�j���E�͊� |
| �R�i���S |
|
1 |
|
|
|
�͊� |
| �R�i���T |
|
1 |
|
|
|
�͊� |
| �R�i���U |
|
|
1 |
|
|
�͊� |
| �R�i���V |
|
|
|
2 |
|
���� |
| �R�i���W |
|
|
|
1 |
|
�j���E�͊� |
| �R�i���X |
|
|
|
|
1 |
|
| �R�i��10 |
|
|
|
|
2 |
|
| �V���J�V�P |
1 |
|
|
1 |
1 |
|
| �V���J�V�Q |
1 |
3 |
|
2 |
3 |
|
| �V���J�V�R |
3 |
|
|
|
|
|
| �V���J�V�S |
|
1 |
|
|
|
|
| �V���J�V�T |
|
|
|
2 |
1 |
|
���̃e�[�}�͂ƂĂ������[���B
�i�������������Ȃ��Ȃ�j���i�̏W�ł��Ȃ��Ȃ�j
�Ƃ����P���Ȑ}���ł͌��Ȃ��B
�Ȃ��Ȃ�A���t�̌͊��͗����͂�̑O�i�K�ł�����A���N��ɂ͔������Ƃ��Đ��Ԍn�̏d�v�Ȗ�����S��
���ƂɂȂ�B�܂��A����߂��ꂪ�h��Ŋ����P��Ă���قǕ��Q�ɂ���ē|�₷�����A������������
�������ƂȂ�B
�L�͂Ȗ����̂����ƁA�̏W�ҐS���Ƃ��Ă͂��Ȃ藎������ł��܂����̂����A��Âɍl����Ɛ芔����
�c���Ă���A�u���H���v�̃q���^�ɂƂ��Ă͊i�D�̎Y���ꏊ����邱�Ƃɂ��Ȃ낤�B
�֑������A������̎G�ؗтɂ�����G��X�V�͍������田�̂��邽�߃m�R��q���^�ȂǂɂƂ��Ă͍D�s��
�ł��邪�A�Ǘ����s���͂��Ă���قǗ����͂ꂪ���Ȃ��Ȃ�A�I�I�N���̐����ɂ͂��s�����������̂ł�
�Ȃ����낤���B
�v�́A�u���R�ȃ|�C���g���Łv�Ɓu���R�ȃ|�C���g�����v�̋ύt�i�g�[�^���o�����X�j���ۂ���Ă��邱�Ƃ��A����
���̑����ɂƂ��ĕs�����B
�G�ؗъǗ��̈�Ƃ��Ă̔��̂�|��A�͎����i�s����T��ŐV���Ȏ������t���o���n�߁A���N�Ԃ���
�ĐV���Ȏ��猞�ꂪ��������Ă����B���������V��ӂ��\�ɂȂ�u�Œ���̒P�ʁv�Ƃ͉����H
�G�ؗтł͂Ȃ��A�ꌩ���̂悤�ɃN�k�M�����邾���ł��A�������Ƃ��ēK���ꍇ������B
�ɒ[�Ȃ��Ƃ������A���t���o�������������{�����Ȃ��Ă��A���̍������������ƂȂ�ݑ���d�˂�
���Ƃ��\�ł��낤�B
�L��ȎG�ؗтł̓q���^�����܂�̂ꂸ�A�ӊO�ɂ��Z��n�̕n��ȗΒn�ō̂��E�E�E�Ƃ����b���悭���ɂ���B
�O�҂ł̓|�C���g���i��ɂ�����҂͂��̋t�A�Ƃ������R���傫�����낤���A�u�K�v�ŏ����̐������Ƃ́H�v
�Ƃ����^��ɑ���̂ЂƂƂ��ċ����[���B
�y�g�s�b�N�ҁz
�i�쐬���j
